Back to
research
Projects J.A.M. van Gisbergen
1. Role of top-down signals in visual motion
detection
2.
Dynamic perceptual updating by vestibular signals
3. Central processing of
vestibular signals for spatial orientation
4. Orientation constancy in
spatial perception
5. Interactions between
control signals of visual and vestibular origin in the generation of rapid eye
movements.
6. Neural mechanisms for the
control of binocular eye movements in direction and depth.
7. Gaze control by eye-head
coordination
1. Role of top-down signals in visual motion detection
Research group: Maaike de Vrijer, Pieter Medendorp en Jan van Gisbergen
Introduction
Detecting
true object motion during self-motion, although seemingly effortless, has to overcome
two nontrivial problems. First, integrate the local motion information derived
from an early fine-grain analysis (the motion seen through many ‘peepholes’)
into a global motion pattern. Second, distinguish object motion from the
effects of self-motion. So far, these problems have been seen as separable and
amenable to sequential analysis. Here, we propose that it would be more optimal
to solve them in conjunction. To evaluate this idea, we propose novel
experiments exploring the conditions for the detection of true object motion
during self-motion.
Report
The
project has just started.
2 Dynamic perceptual updating by vestibular signals
Research group: Rens Vingerhoets, Pieter Medendorp
and Jan Van Gisbergen
Introduction
Major issues in the study of spatial orientation concern the questions of how
the brain detects self movement and how this information is used to maintain a
stable percept of external space. In
this project, perceptual stability will be tested on a moment to moment basis
while the subject is continuously rotated about a tilted (off-vertical) axis.
In the absence of visual cues, the brain depends strongly on vestibular
information to reconstruct head rotation and head position in space. The
semicircular canals signal head rotation but they only detect angular
acceleration so that their signal dies out as rotation continues. The otoliths
detect linear acceleration forces but they cannot distinguish between the pull
of gravity and the effect of translation. Interpretation of their ambiguous
signal requires integration of available rotation signals by an internal model
in the brain. When these body rotation signals are reliable, as during rotation
in the light, percepts of body movement are roughly veridical. However, during
fast off-vertical rotation in the dark (i.e., about a tilted axis), subjects
have illusory translation and tilt percepts that do not correspond to the
actual pure rotation stimulus. This combination of illusory effects has been
interpreted in terms of incorrect otolith signal disambiguation in conditions
where rotation information from the canals is misleading. To really test this
canal-otolith interaction hypothesis, quantitative perceptual data at high
temporal resolution are essential. The project is designed to provide such data
for the first time. The major objective in collecting these data is to test to
what extent current dynamic orientation models, which have been based and
tested mainly on oculomotor data, can also be applied to perception.
Report
Human spatial orientation relies on vision, somatosensory cues and
signals from the semicircular canals and the otoliths. The canals measure
rotation, while the otoliths are linear accelerometers, sensitive to tilt and
translation. To disambiguate the otolith signal, two main hypotheses have been
proposed: frequency segregation and canal-otolith interaction. So far these
models were based mainly on oculomotor behavior. In this study we investigated
their applicability to human self-motion perception. Six subjects were rotated in yaw
about an off-vertical axis (OVAR) at various speeds and tilt angles, in
darkness. During the rotation, subjects indicated at regular intervals whether
a briefly presented dot moved faster or slower than their perceived selfmotion.
Based on
such responses, we determined the time course of the selfmotion percept and characterized its steady-state by a
psychometric function. The psychophysical results were consistent with anecdotal
reports. All subjects initially sensed rotation, but then gradually developed a
percept of being translated along a cone. The rotation percept could be
described by a decaying exponential with a time constant of about 20 s.
Translation percept magnitude typically followed a delayed increasing
exponential with delays up to 50 s and a time constant of about 15s. The
asymptotic magnitude of perceived translation increased with rotation speed and
tilt angle, but never exceeded 14 cm/s. These results were most consistent with
predictions of the canal-otolith interaction model, but required parameter
values that differed from the original proposal. We conclude that canal-otolith
interaction is an important governing principle for self-motion perception that
can be deployed flexibly, dependent on stimulus conditions.
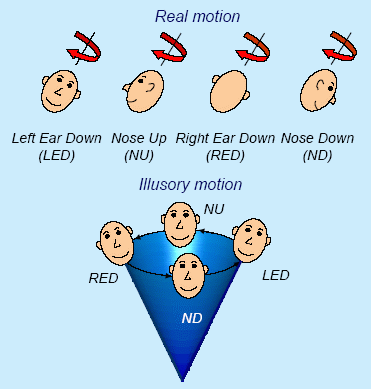
Rotation about an axis tilted relative to
gravity generates striking illusory motion percepts. As the initial percept
of body rotation gradually subsides, a feeling of being translated along a
cone emerges. The direction of the illusory conical motion is opposite to
the actual rotation.
Recent publications
·
Vingerhoets, R.A.A. , Medendorp, W.P.
and Gisbergen J.A.M. van
Time course and magnitude of illusory translation perception during off-vertical
axis rotation
J. Neurophysiol. 95:
1571-1587, 2006 (pdf)
- Vingerhoets, R.A.A., van Gisbergen,
J.A.M. and Medendorp, W.P.
Verticality perception during off-vertical axis rotation.
J. Neurophysiol. 97, 3256-3268, 2007 (pdf)
3. Central processing of
vestibular signals for spatial
orientation
Research group: Ronald Kaptein and Jan Van Gisbergen

Introduction
The vestibular system is important for the detection of the direction of
gravity and for the analysis of body movements, which generally consist of
various combinations of rotations and translations. For this purpose, it has
specialized sensors (canals and otoliths), but limitations in their coding
properties complicate the analysis. The canal afferents are insensitive to low
rotation frequencies. The otoliths, for elementary physical reasons, cannot
distinguish between the inertial forces caused by translation and those exerted
by tilt relative to gravity. Yet, this tilt/translation distinction is vital
for survival. The problem how the brain achieves otolith disambiguation, a hot
topic surrounded by controversy, is the central theme of this proposal. The
project will investigate the consequences of shortcomings in the disambiguation
process for spatial orientation and spatial localization.
Report
Using vestibular sensors to maintain visual stability during changes in head
tilt, crucial when panoramic cues are not available, presents a computational
challenge. Reliance on the otoliths requires a neural strategy for resolving
their tilt/translation ambiguity, such as canal-otolith interaction or
frequency segregation. The canal signal is subject to bandwidth limitations. In
this study, we assessed the relative contribution of canal and otolith signals
and investigated how they might be processed and combined. The experimental
approach was to explore conditions with and without otolith contributions in a
frequency range with various degrees of canal activation.
We tested the perceptual stability
of visual line orientation in six human subjects during passive sinusoidal roll
tilt in the dark at frequencies from 0.05 to 0.4 Hz (30 deg peak-to-peak). Since subjects were
constantly monitoring spatial motion of a visual line in the frontal plane, the
paradigm required moment-to-moment updating for ongoing ego motion. Their task
was to judge the total spatial sway of the line when it rotated sinusoidally at
various amplitudes. From the responses we determined how the line had to be
rotated to be perceived as stable in space. Tests were taken both with (subject
upright) and without (subject supine) gravity cues.
Analysis of these data showed that
the compensation for body rotation in the computation of line orientation in
space, while always incomplete, depended on vestibular rotation frequency and
on the availability of gravity cues. In the supine condition, the compensation
for ego motion showed a steep increase with frequency, compatible with an
integrated canal signal. The improvement of performance in upright, afforded by
graviceptive cues from the otoliths, showed low-pass characteristics.
Simulations showed that a linear combination of an integrated canal signal and
a gravity-based signal can account for these results.
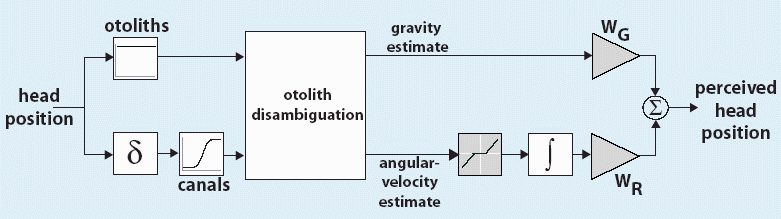
Linear summation of graviceptive and rotational
cues.
First,
separate estimates of the direction of gravity and angular velocity are obtained.
These are weighted and added to obtain the estimate of head rotation. Two
disambiguation strategies, frequency segregation and canal-otolith interaction,
were simulated.
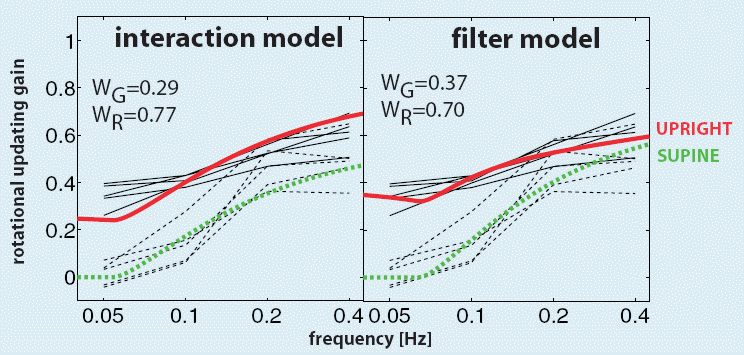
Model fits.
Both versions of the model can
mimic the high-pass behavior in both conditions. The filter model, relying on
frequency segregation for disambiguating the otolith signal, reproduces the
low-pass characteristics of the otolith signal most accurately. To explain the
data, both models require strong canal contribution.
Recent publications:
Kaptein, R.G.,
Gisbergen, J.A.M. van
Interpretation
of a Discontinuity in the Sense of Verticality at Large Body Tilt.
J Neurophysiol 91: 2205-2214, 2004
Kaptein, R.G. and
Gisbergen, J.A.M. van
Nature
of the transition between two modes of external space perception in tilted
subjects.
J. Neurophysiol. 93: 3356-3369, 2005
Kaptein, R.G. and
Gisbergen, J.A.M. van
Canal
and otolith contributions to visual orientation constancy during sinusoidal
roll rotation
J. Neurophysiol 95: 1936-1948, 2006 (pdf)
Kaptein,
R.G.
Vestibular
Contributions to visual stability
PhD thesis, Radboud University Nijmegen, 2006 (pdf)
4. Orientation constancy
in spatial perception
Research group: Anton van Beuzekom and Jan van
Gisbergen
Introduction
One of the key questions in spatial perception is whether the brain has a
common representation of gravity that is generally accessible for various
perceptual orientation tasks. To evaluate this idea, we compared the ability of
passively and actively-tilted subjects to indicate earth-centric directions in
the dark with a visual and an oculomotor paradigm and to estimate their body
tilt relative to gravity.
Report We compared the ability of
passively-tilted subjects to indicate earth-centric directions in the dark with
a visual and an oculomotor paradigm and to estimate their body tilt relative to
gravity. Subjective earth-horizontal and earth-vertical data were collected,
either by adjusting a visual line or by making saccades. In all spatial
orientation tests, whether involving space-perception or body-tilt judgments,
passively-tilted subjects made considerable systematic errors which mostly
betrayed tilt underestimation (Aubert-effect) and peaked near 130 deg tilt.
However, the Aubert-effect was much smaller in body tilt estimates than in
spatial oculomotor pointing, implying that the underlying signal processing
must have been different. Furthermore, in actively-tilted subjects, the body
tilt estimates became much better but space perception performance was
virtually identical to the passive tilt condition.
These findings are discussed in the context of a conceptual model in an attempt
to explain how the different patterns of systematic and random errors in
external-space and self-tilt perception may come about.
Recent publications
Van
Beuzekom AD, and Van Gisbergen JAM (1999)
Comparison of tilt estimates based on line settings, saccadic pointing and
verbal reports.
Ann N Y Acad Sci 871: 451-454.
Van Beuzekom AD, and Van Gisbergen JAM (2000)
Properties
of the internal representation of gravity inferred from spatial direction and
body tilt estimates.
J Neurophysiol 84: 11-27.
Van Beuzekom AD, Medendorp WP, and Van Gisbergen JAM (2001)
The subjective vertical and the sense of self
orientation during active body tilt.
Vision Research 41: 3229-3242.
5. Interactions between
control signals of visual and vestibular origin in the generation of rapid eye
movements.
Research group: Anton van Beuzekom and Jan Van Gisbergen
Introduction
Rapid eye movements to different sensory inputs share a common final pathway.
There is clear evidence that the pathways for saccades to visual, auditory and
tactile targets have already converged at the level of the superior colliculus
(SC) where movement vectors are coded spatially in a motor map. It is not clear
whether this applies also to vestibularly induced rapid eye movements (quick
phases). Neural activity related to quick phases has been demonstrated in the
SC but its functional significance remains uncertain. Another possibility to be
considered is that quick phase signals enter the saccadic pathway at a more
peripheral stage where signals are coded temporally, separately for each
component, i.e. at the level of burstcells. Accordingly, there are at least two
levels of possible interaction between visual and vestibular signals for the
generation of rapid eye movements (vector and component stage). When both
visual and vestibular inputs impinge on the system, interesting interactions
occur whose precise nature and neural basis have hardly been studied. This
project was designed to study these interactions behaviorally and neurophysiologically.
Our aim was to elucidate how the SC participates in the control of vestibularly
induced rapid eye movements and to clarify to what extent this role and the
interaction findings can be reconciled with its role in foveation.
Report To investigate these issues, we
have studied in the monkey to what extent saccades induced by electrical
microstimulation in the SC (E-saccades) can be modified by concurrent
sinusoidal rotation about a vertical axis. We found clear kinematic effects
which cannot readily be understood by interactions at this level. Vectorial
peak velocity in E-saccades of a given amplitude increased with head velocity
for rotations into the half field where the saccade was directed and decreased
for rotation in the opposite direction. Interestingly, this effect was due to a
specific effect on the horizontal component; the vertical component did not
show systematic kinematic changes. We suggest that the kinematic effect may be
due to preparatory quick-phase signals, impinging at the burstcell level, that
become expressed once the pause cell gate has been opened by collicular
signals. If interaction at a collicular level were responsible for the
kinematic effects, one would expect comparable effects in both components. To
explore the nature of visuo-vestibular interactions further, we have
investigated how the human saccadic system copes with the interfering effects
of ongoing horizontal nystagmus when attempting to generate oblique prosaccades
or antisaccades. The results suggest that voluntary and reflexive rapid eye
movements can be programmed in parallel. The outcome of this process shows a
degree of independence between horizontal and vertical components which is not
readily understandable in terms of interactions at a vectorial coding stage
such as in the collicular map.
Recent publications
Van
Beuzekom AD, and Van Gisbergen JAM (2002)
Collicular
microstimulation during passive rotation does not generate fixed gaze shifts.
J Neurophysiol , 87, 2946-2963, 2002.
Van Beuzekom AD, and Van Gisbergen JAM (2002)
Interaction
between visual and vestibular signals for the control of rapid eye movements.
J Neurophysiol., 88, 306-322, 2002.
6. Neural mechanisms for
the control of binocular eye movements in direction and depth.
Research group: Vivek Chaturvedi and Jan Van
Gisbergen
Introduction
It is generally assumed that, at a peripheral level, two distinct oculomotor
control systems are involved in the execution of binocular gaze shifts in three
dimensions. When the target movement calls only for a change in gaze direction,
the movement is controlled by the fast saccadic system. Target changes in depth
elicit slow vergence movements. When both subsystems are called into action by
a target step in 3D (direction and depth component) they do not act
independently: Their responses show non-additive dynamic behaviour in that the
vergence component is now clearly faster. The aim of this project was to
investigate various aspects of the coupling between the two systems relying on
both behavioural analysis and on neurophysiological studies.
Report We investigated the role of the
monkey superior colliculus (SC) in the control of combined saccade-vergence
movements by assessing the perturbing effects of microstimulation. We elicited
an electrical saccade by stimulation while the monkey was preparing a visually-guided
movement to a near visual target. We showed that that artificial intervention
in the SC, while a 3D refixation is prepared or is ongoing, can effect the
timing (WHEN) and the metric-specification (WHERE) of both saccades and
vergence. This effect would be expected if the population of movement cells at
each SC site is tuned in 3D, combining the well-known topographical code for
direction and amplitude with a nontopographical depth representation. For an
eye movement to a near target, stimulation at a rostral site leds to a
significant suppression of the pure vergence response during the period of
stimulation. When these paradigms were implemented for 3D refixations, the
saccade was inactivated, as expected, while the vergence component was often
suppressed more than in the case of the pure vergence. We conclude that the
rostral SC, presumably indirectly via connections with the pause neurons, can
affect vergence control for both pure vergence and combined 3D responses..
Recent publications
Chaturvedi
V, and Van Gisbergen JAM (1998)
Shared
target selection for combined version-vergence eye movements.
J Neurophysiol 80: 849-862
Chaturvedi, V. and Van Gisbergen, J.A.M. (1999)
Perturbation
of combined saccade-vergence movements by microstimulation in monkey superior
colliculus.
J. Neurophysiol. 81: 2279-2296.
Chaturvedi, V. and Van Gisbergen, J.A.M.
Stimulation
in the rostral pole of monkey superior colliculus: effects on vergence eye
movements.
Exp. Brain Res. 132:72-78, 2000.
7. Gaze control by
eye-head coordination
Research group: J.A.M. van Gisbergen, C.C.A.M.
Gielen
Introduction
Fixating a visual target during head movements requires specific eye movements
in order to compensate for movements of the eyes relative to the target.
Several sensory systems (e.g. semi-circular canals, otolith system, visual
system, muscle spindles in neck muscles) contribute to the accurate
stabilization of gaze on targets in 3-D space. The aim of this project is 1) to
investigate the characteristics of eye movements in gaze stabilization in
natural head movements and 2) to determine the role of various sensory systems
to gaze stabilization.
Report
We have investigated gain modulation (context compensation) of the
vestibulo-ocular reflex (VOR) for binocular gaze stabilization in human
subjects during voluntary yaw and pitch head rotations. Movements of each eye
were recorded both when attempting to maintain gaze on a small visual target in
a darkened room and after its disappearance (remembered target).
Linear regression analysis
on the version gain as a function of target distance yielded a slope representing
the influence of target proximity and an intercept corresponding to the
response at zero vergence (default gain). The slope of the fitted relationship,
divided by the geometrically-required slope provided a measure for the quality
of version context compensation ('context gain'). In both yaw and pitch
experiments, we found default version gains close to one even for the
remembered target condition, indicating that the active VOR for far targets is
already close to ideal without visual support. In near target experiments, the
presence of visual feedback yielded near unity context gains, indicating close
to optimal performance. For remembered targets, the context gain deteriorated
but was still superior to performance in corresponding passive studies reported
in the literature.
We examined how frequency
and target distance, estimated from the vergence angle, affected the
sensitivity and phase of the version component of the translational VOR (tVOR)
and compared the results to the requirements for ideal performance. Linear
regression analysis of the version-sensitivity relationship yielded a slope
representing the influence of target distance. The ratio of this slope and the
slope required for ideal stabilization provided a measure for the degree of 'distance
compensation' in the tVOR. The results show that distance compensation behaves
according to low-pass characteristics in each target condition. It declined
from 1.00 to 0.84 for visual targets and from 0.87 tot0.57 for remembered
targets in the frequency range 0.25 - 1.5 Hz. The intercept obtained from the
regression yielded the tVOR response at zero vergence and specified the
'default sensitivity' of the tVOR. Default sensitivity increased with frequency
from 0.02 to 0.10 deg/cm for visual targets and from 0.04 to 0.16 deg/cm in
darkness. The phase delays of version angular eye velocity relative to
translational eye velocity increased on average from 2 deg to 7 deg. In
comparison with earlier passive studies, the active tVOR in the dark performs
much better at all frequencies where comparison was possible. We conclude that
an additional nonvestibular signal with low-pass characteristics contributed to
the tVOR during self-generated head translations.
Recent publications
W.P.
Medendorp, J.D. Crawford, D.Y.P. Henriques, J.A.M. van Gisbergen, C.C.A.M.
Gielen
Kinematic
strategies for upper arm-forearm coordination in three dimensions.
J. Neurophysiol. 84, 2302-2316, 2000.
W.P. Medendorp, J.A.M. van Gisbergen, S. van Pelt, C.C.A.M. Gielen
Context
compensation in the vestibulo-ocular reflex during active head rotations.
J. Neurophysiol. 84, 2904-2917, 2000.
W.P. Medendorp, J.A.M. van Gisbergen, C.C.A.M. Gielen
Human gaze
stabilization during active head translations.
J. Neurophysiol. 87, 295-304, 2002.