PROJECTS
IN THE LAB OF JEROEN GOOSSENS:
1) Quantitative assessment of slow vision*
2) Origin of slow vision: Contributions of the central and peripheral nervous system*
3) The effect of bifocals in children with Down Syndrome*
4) Perceptual learning to reduce crowding effects in visually impaired children*
5) Decision-making under uncertainty and ambiguity*
6) Interactions between binocular rivalry and saccades
7) The role of lateral inhibition in binocular motion rivalry
8) Influence of stimulus strength in binocular motion rivalry
9) The topography of egomotion processing*
10) Representation of head-centric visual flow in the human motion complex
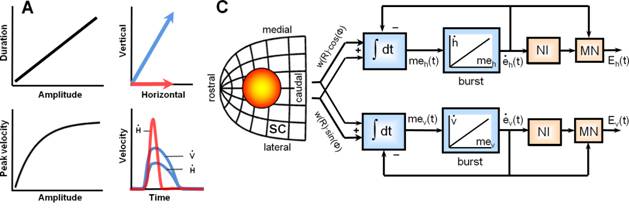
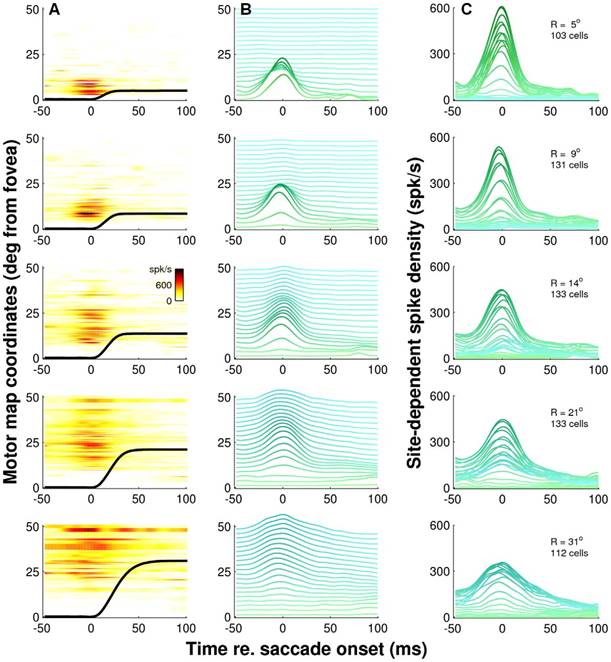
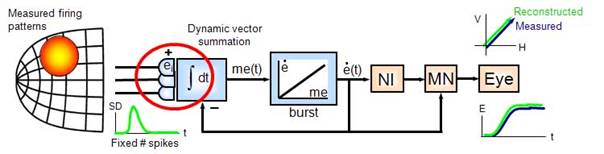
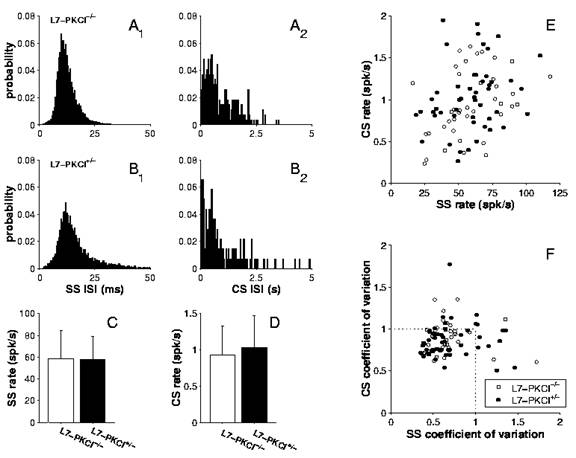
11) Optimal control of saccades by the monkey superior colliculus
12) Effect of reflex blinks on saccade perturbations in humans
13) Dynamic ensemble coding of saccades in monkey superior colliculus
14) Motion processing in human extra-striate visual cortex
15) Processing of egocentric and allocentric spatial visual information
16) Plasticity of compensatory eye movements
17) Eye head coordination in 2D under different sensorimotor conditions
18) see also
*recently started projects or work in progress
1.
quantitative assesment of
slow vision
Research group: J. Goossens, A. Barsingerhorn and N. Boonstra (Bartiméus)
Summary
For the majority of
tasks in daily life we rely on visual information; the demands on our visual
system become even greater in dynamic situations, for example in traffic, where
visual information changes rapidly over time.
Most of young adults and children are able to deal with this information
efficiently. However, if it takes too long to process and respond to visual
stimuli problems occur. Existing tests
of visual acuity do not take into account the time that is needed to respond.
In the Netherlands, people who seek help because they encounter problems of
slow vision do not receive assistance from institutes for the visually impaired
or rehabilitation facilities. At present the definition of visual impairment is
primarily based on distant visual acuity and visual field. There are no
internationally accepted tests to measure reaction time in acuity testing.
Nevertheless, many children and adults, both normally sighted as visually
impaired, complain about problems in coping with various daily activities due
to slow vision. For example, children need more time to complete exercises at
school and both adults as well as children encounter problems when using
digitized information (e.g. using computers) or with participation in traffic. We need validated tests to measure
the time children and adults need to respond to visual stimuli. Reaction times
measurements could be combined with the measurement of visual acuity (at
distance and near). With this method we can then estimate the delay that
normally sighted and visually impaired persons experience in daily activities
such as work, school and traffic.
Because visual task performance is
characterized by speed-accuracy tradeoffs, measurements of both speed and
accuracy are key to better quantify visual impairment. In this study, we will
therefore develop new tools to measure “perception time” during the assessment
of visual acuity. In principle our perception-time measurements could depend on
several factors including: 1) the time subjects need to foveate the optotype by
making eye movements towards it, 2) the stability with which they can keep
their eyes fixated on the optotype, and 3) the time they need to identify a
foveated optotype. Little is known, however, about the way children develop
strategies to focus on a distant target, how they use eye movements (saccades)
and how they cope with disturbing factors (distracters, noise, crowding). To further assess the contribution of these
factors, we will therefore measure the eye movements during the modified
acuity-speed tests using a new stereoscopic eye-tracking instrument that works
without individual calibration.
Funded by: ODAS
foundation
Back to top, to Jeroen's Homepage, or to the Department’s homepage
2.
neural origin of slow vision:
contributions of the central and peripheral nervous system
Research group: J. Goossens, K. Woutersen, A.V. van den Berg , N. Boonstra (Bartiméus) and T. Theelen (Ophthalmology dept)
Summary
A group of children and adults has presented itself to
the Bartiméus institute and the department of ophthalmology Radboudumc with
complaints about slow vision at school or work but they do not meet any of the
diagnostic criteria for visual impairment. Indeed, task performance in daily
life is more complex than visual
performance alone since it involves motor and cognitive factors as well.
In this project, we will first determine if our newly
developed speed-acuity test or the existing useful field of view (UFOV) test
are valid psychophysical methods to better characterize the impairment in this
group of patients. Because the origin of their problem is unknown, we will also
investigate their visual and attentional system using a variety of techniques.
Both peripheral and central factors may contribute. In collaboration with the ophthalmology
department we will therefore test
the integrity of the retina using functional optical coherence tomography (fOCT)
and electrophysiological examination (visual-evoked potentials (VEP) and
(multifocal)electroretinogram (ERG/mERG)). To assess the contribution of more
central (e.g., attention) deficits, we will measure fMRI and MEG activity. By linking the neuroimaging results to the psychophysical results we hope to gain
a much better understanding of the neural dysfunctioning in slow vision.
Funded by: Donders
Centre for Neuroscience of the Donders
Institute
Back to top, to Jeroen's Homepage, or to the Department’s homepage
3.
The effect of bifocals in
children with down syndrome
Research group: J.
Goossens, C. de Weger-Zijlstra and N. Boonstra (Bartiméus)
Summary
DS is one of the most common genetic anomalies,
occurring in about 14.6 in 10000 live births in the Netherlands in 2007. Near
vision is reduced in 86 to 100% of the children with Down Syndrome (DS). This
is an additional barrier achieving their maximum potential in development. In
the last two decades many research is done to find out the differences in
ocular findings between children with and children without DS. Refraction
errors, which have to be corrected with glasses, are common (percentages vary
from 40 to 90%) and aggravate over time. The accommodation (focussing for near)
is consistent reduced in 50 to 100% of children with DS and does not improve
with age. In contrast to children without DS glasses for distance vision don’t
improve near vision. Children with DS see
blurred at near. Some authors have suggested a relationship between this
blurred retinal image and the absence of emmetropisation (decrease of
refraction error), others see a crucial relationship with the defective visual
development of children with DS, presenting in visual acuities that do not
reach normal levels, generally 20/40 or lower. Moreover the effort to
accommodate may give rise to strabismus, which occurs far more often in
children with DS, in 15 to 47% (versus 3 to 4% in normal population) and could
be avoided or cured by wearing the right glasses. Bifocal correction is such a
tailor-made treatment for the eye disorders in DS. Results shown in smaller
studies on the effect of bifocals are encouraging: 1) Improved
visual acuity for near, 2) Significantly more accurate accommodation in the
bifocal-treatment group, 3) Positive impact on visual functioning: Faster and
improved performance on visual perceptual and some early literacy skills. 4)
Bifocals were used with good compliance, and 5) Prevention of avoidable visual
impairment. In this project, we will perform a multicentre randomised
controlled trail (RCT), the most suitable design for effects of interventions.
Our objectives are 1) to generate evidence of improved visual functioning (both
visual acuity at near and at distance as well as accommodation accuracy) with
bifocals and 2) to assess task-readiness by monitoring executive functions.
Funded by: LSBS,
Oogfonds, ODAS and Novartis
Back to top, to Jeroen's Homepage, or to the Department’s homepage
4.
Perceptual learning to reduce
crowding effects in visually impaired children
Research group: J.
Goossens, B. Huurneman and N. Boonstra (Bartiméus)
Summary
Foveal crowding is a phenomenon in which adjacent
objects degrade recognition of an object in central vision. A higher degree of
crowding has been associated with slower reading rates and could also hinder
the acquisition of reading skills. Visually impaired children show a higher
degree of crowding than children with normal vision. Classically, crowding has
been explained by three causes: contour interaction (sensory aspect),
fixational stability (motor aspect), and selective attention (cognitive
aspect). However, the unique contribution of these individual factors on
explaining the strength of crowding effects has not been investigated yet.
A recent therapy to decrease crowding is based on
Perceptual Learning (PL). PL occurs when visual tasks are practiced repeatedly,
and can lead to drastic, long-term improvements in performing these tasks. PL
can improve visual acuity, contrast sensitivity and spatial resolution of the
visual system in children with amblyopia. It can reduce foveal and peripheral
crowding in students with normal vision, adults with normal vision, and adults
with amblyopia. PL in adults with low vision can improve visual search speed
and accuracy and reading speed. PL exercises are usually presented on a
computer. This has several advantages: it can be performed at home, progress
can be easily tracked, and task difficulty is automatically adjusted to the
child’s abilities.
The objectives of this study are: 1) to
disentangle the contribution of different aspects on the strength of crowding
effects in visually impaired children, and 2) to evaluate the effect of PL on
the reduction of crowding effects in visually impaired children. More
fundamental research questions that we will address are: Can oculomotor control
be improved with PL? Or are training effects only observable in the sensory
domain? Are task improvements after PL specific (only training task) or are
there general improvements in oculomotor control, visual acuity, or reading
performance?
Funded by: ODAS, LSBS and Vereniging Bartiméus-Sonneheerdt
Back to top, to Jeroen's Homepage, or to the Department’s homepage
5.
Decision-making under
uncertainty and ambiguity
Research group: J.
Goossens, J. Kalisvaart and I. Klaver
Summary
When confronted with
an ambiguous stimulus, the brain somehow has to decide which of the two
possible images is perceived, making binocular rivalry and visual
decision-making conceptually very similar processes. Yet, in the literature,
these two phenomena have been explained by distinctly different mechanisms and
models. In a human psychophysical study (Kalisvaart et al., 2011), we therefore
studied decision-making under rivalrous and noisy stimulus conditions using
random-dot motion stimuli that differed between the left and the right eye in
different ways. We found that speed and accuracy of motion-direction discrimination
was similar when observers viewed noisy, unambiguous motion patterns in which
signal dots were either at identical or at different, uncorrelated locations
for the two eyes. This result is consistent with a race between two monocular
drift-diffusion processes. However, reaction times increased significantly
under rivalry conditions and this increase could not be explained by motion
transparency. Our data thus revealed competitive binocular interactions. We
developed a model which combines the key elements of diffusion models (i.e.,
accumulation of evidence) and rivalry models (i.e., feedback competitive
interactions) and showed that our new model is able to predict the choice
behavior and reaction times in both an unambiguous but noisy decision making
task, as well as in a situation with ambiguous motion.
We also tested one monkey subject under similar task conditions while
recording single-unit activity in the posterior parietal cortex (PPC), and we
are currently collecting data from a second animal. The behavioral data
indicate that their motion judgments (which they indicate by manual lever
presses) under uncertainty and rivalry is comparable to that of humans, and
preliminary analysis of the cell data indicates that response-delay in the rivalry
condition is reflected in the activity of PPC neurons.
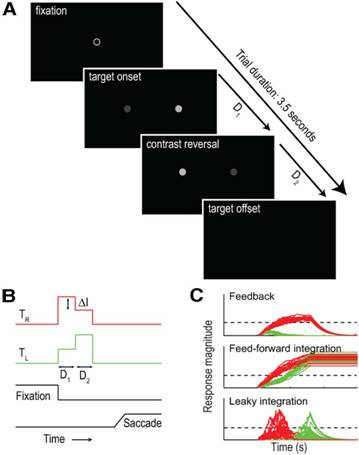
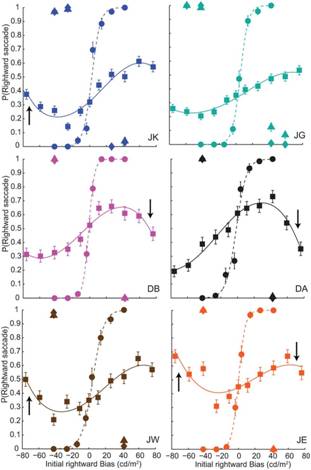
In another human psychophysical study (Kalisvaart et al., 2013) we
studied saccade target-selection. In the ambiguous condition we presented two
simultaneously-flashed targets that reversed their intensity difference during
each presentation, and instructed subjects to make a saccade towards the
brightest target. All subjects preferentially chose the target that was
brightest during the first stimulus phase. Unless this first phase lasted only
40 ms, this primacy effect persisted even if the second, reversed-intensity
phase lasted longer. This effect did not result from premature commitment to
the initially-dominant target; a strong target imbalance in the opposite
direction later on drove nearly all responses towards that location. Moreover,
there was a non-monotonic relation between primacy and target imbalance;
increasing the target imbalance beyond 40 cd/m2 caused an
attenuation of primacy. These are the hallmarks of hysteresis, predicted by
models in which target-representations compete through strong feedback. Our
findings thus showed that our feedback competition model, designed originally
to describe the choice behavior at the onset of binocular rivalry could also
predict our subjects’ saccade behavior. Ongoing experiments aim to test this
feedback competition model using neurophysiological recordings in rhesus
macaques.
|
Intensity reversal
paradigm. See Kalisvaart et. al., J. Neurosci., 2013 for details |
Choices
are predicted by models in which target representations compete through strong
feedback |
Publications
o
Kalisvaart
J., Noest A.J., van den Berg A., Goossens J.
Saccade target selection relies on feedback competitive signal integration.
J. Neurosci., 33:12077-12089, 2013
o Kalisvaart J., Klaver I., Goossens J.
Motion discrimination under uncertainty and ambiguity.
J. Vision, 11:1-21, 2011
Funded by: VIDI Innovational Research Incentives Scheme of NWO Earth and Life Sciences
Back to top, to Jeroen's Homepage, or to the Department’s homepage
6.
Interactions between
binocular rivarly and saccades
Research group: J.
Goossens, J. Kalisvaart,
Summary
A saccade moves the
image over the retina. Because of the retinotopic organization of the visual
system, after the saccade a different cell population is stimulated by the
stimulus than before. This new population clearly has a different adaptation
history. Indeed, there is quite some evidence showing that there is some
interaction between eye movements and perceptual alternations in binocular
rivalry. Several studies, however, have shown by either compensating for
occurring eye movements or using afterimages that eye movements are not
necessary for perceptual alternations to occur. Thus, the effects of saccades
on binocular rivalry are not trivial and studying saccades is an interesting
way to reveal the location of the rivalry process in the brain. If rivalry is
purely a low level, local process, the new population of cells that gets
stimulated after the saccade would see the stimulus after the saccade as a new
stimulus, resulting in onset rivalry that is indistinguishable from the
situation in which the stimulus moved, instead of the eyes. In contrast, if
higher order processes are involved in binocular rivalry, the percept after a
saccade might be different from the percept after a passive stimulus movement.
We therefore investigated the effect of saccades on binocular rivalry. In one
set of experiments (Kalisvaart et al., PLoS ONE, 2011), we studied the effect
of a single, 4° saccade on the rivalry process in comparison with a similar but
passive movement of the stimulus over the retina. We found that both active
saccades and passive stimulus movements trigger onset rivalry, but not to the
same extent, suggesting the involvement of extra-retinal signals (eye-movement
related) in binocular rivalry.
In a second series of experiments (Kalisvaart & Goossens, PLoS ONE,
2013), we instructed subjects to make many saccades of different sizes while
watching an ambiguous stimulus, and compared the perceptual switch probability
around the time of the saccade with the behavior around similar, but passively
induced, stimulus movements. We found a strong relation between large (>1°)
saccades and perceptual switches, but small (<1°) saccades did not seem to
influence the rivalry process at all. Stimulus jumps of all amplitudes, on the
other hand, were found to be related to perceptual switches. These results
further corroborate our conclusion that extra-retinal signals are involved in
the rivalry process.
Interestingly, in the macaque monkey,
we find that during sustained binocular motion rivalry many cells in PPC
exhibit a burst of activity right after a saccade. We suspect that these are
visual transients evoked by the eye displacement. We are now pursuing the
question whether these responses are different for different saccade
amplitudes, and whether the occurrence of these bursts is linked to the
observed correlation between percept switches and saccade occurrences.
Publications
o
Kalisvaart J., Goossens J.
Influence of retinal image shifts and extra-retinal eye movement signals
on binocular rivalry alternations.
PLoS ONE, 8(4):e61702, 2013
o
Kalisvaart J., Rampersad S., Goossens
J.
Binocular onset rivalry at the time of saccades
and stimulus jumps.
PLoS ONE, 6(6):e20017, 2011
Funded by: VIDI Innovational
Research Incentives Scheme of NWO Earth
and Life Sciences
Back
to top, to Jeroen's Homepage,
or to the Department’s homepage
7.
The Role of lateral
inhibition in binocular motion rivalry
Research group: J.
Goossens and A. Platonov
Summary
It is generally believed that percept alternations in binocular rivalry
result from the interplay between mutual inhibition and slow adaptation of the
competing percepts, but the nature of this mutual inhibition is still
unknown. To fill this gap, we presented human subjects with dichoptic
random-dot motion stimuli, and manipulated the angle between the monocular
directions of motion from pure opponent horizontal motion to pure vertical
motion in the same direction (Platonov & Goossens, 2013a). We found that decreasing the angle between the two monocular
directions of motion increased the predominance and mean dominance durations of
the motion pattern presented to the ocular dominant eye (as identified by the
hole-in-card test). This effect was stronger if the contrast of the stimuli was
lowered. Computer simulations showed that these features are a hallmark of
weighted lateral inhibition between populations of directionally-tuned
motion-sensitive neurons. Our findings thus suggest dominance and suppression
in binocular rivalry arises naturally from this fundamental principle in
sensory processing.
|
Parsimonious
population model for binocular motion rivalry. See Platonov & Goossens, J. Vision, 2013, for details. |
Response
of the network for decreasing motion–direction disparity and contrast matches
the behavior of our subjects. |
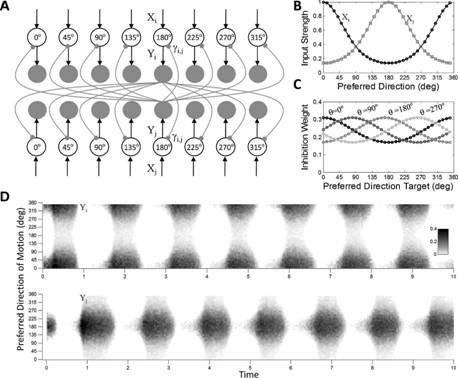

Publications
o
Platonov A., Goossens J.
The role of lateral inhibition in binocular motion rivalry.
J. Vision, doi:10.1167/13.6.12, 2013
Funded by: VIDI Innovational Research Incentives Scheme of NWO Earth and Life Sciences
Back to top, to Jeroen's Homepage, or to the Department’s homepage
8.
Influence of stimulus
strength in binocular motion rivalry
Research group: J.
Goossens and A. Platonov
Summary
Levelt’s four propositions (L1-L4), which characterize the relation
between changes in “stimulus strength” in the two eyes and percept
alternations, are considered as the benchmark for binocular rivalry models. It
was recently demonstrated that adaptation mutual-inhibition models of binocular
rivalry capture L4 only in a limited range of input strengths, predicting an
increase rather than a decrease in dominance durations with increasing stimulus
strength for weak stimuli. This
observation challenges the validity of
those models, but possibly L4 itself is invalid. So far, L1-L4 have been tested mainly by
varying the contrast of static stimuli, but since visual awareness brakes down
at low contrasts, it has been difficult to study L4. To circumvent this
problem, and to test if the recent revision of L2 has more general validity, we
studied changes in binocular rivalry evoked by manipulating coherence of
oppositely-moving random-dot stimuli in the two eyes, and compared them against
the effects of stimulus contrast (Platonov & Goossens, 2013b). Both
contrast and coherence manipulations in one eye produced robust changes in both
eyes; dominance durations of the eye receiving the stronger stimulus increased
while those of the other eye decreased, albeit less steeply. This is
inconsistent with L2 but supports its revision. When coherence was augmented in
both eyes simultaneously, dominance durations first increased at low coherence,
and then decreased for further increases in coherence. The same held true for
the alternation periods. The initial increase in dominance durations was absent
in the contrast experiments, but with coherence manipulations, rivalry could be
tested at much lower stimulus strengths. Thus, we found in humans that L4, like
L2, is only valid in a limited range of stimulus strengths. Outside that range,
the opposite is true. Apparent discrepancies between contrast and coherence
experiments could be fully reconciled with adaptation mutual-inhibition models
using a simple input transfer-function.
Publications
o
Platonov A., Goossens J.
Influence of contrast and coherence on the temporal dynamics of binocular
motion rivalry.
PLoS ONE, 8(8):e71931, doi:10.1371/journal.pone.007193, 2013
Funded by: VIDI Innovational Research Incentives Scheme of NWO Earth and Life Sciences
Back to top, to Jeroen's Homepage, or to the Department’s homepage
9.
The topography of egomotion
perception
Research group: A.V. van den Berg, D. Arnoldussen, J. Goossens
Summary
Recent investigations
indicate that retinal motion is not directly available for perception when
moving around (1), possibly pointing to suppression of retinal speed
sensitivity in motion areas. Here, we investigated the distribution of
retino-centric and headcentric representations of self rotation in human
lower-tier visual motion areas. fMRI responses were measured to a set of visual
self-motion stimuli with different levels of simulated gaze and simulated head
rotation. A parametric GLM-analysis of the BOLD responses revealed sub-regions
of area V3A, V6+, MT, and MST that were specifically modulated by the speed of
the rotational flow relative to the eye and head. Pursuit signals, which link
the two reference frames, were also identified in these areas. To our knowledge,
these results are the first demonstration of multiple visual representations of
self-motion in these areas. The
existence of such adjacent representations points to early transformations of
the reference frame for visual self-motion signals and a topography by visual
reference frame in lower-order motion-sensitive areas. This suggests that visual decisions for
action and perception may take into account retinal and head-centric motion
signals according to task requirements.
Publications
o
Arnoldussen, D.M., Goossens, J., van den Berg, A.
Visual perception of axes of head rotation.
Front. Behav. Neurosci., 7:11. doi: 10.3389/fnbeh.2013.00011, 2013
o
Arnoldussen
D, Goossens J., van den Berg A.
Adjacent visual representations of self-motion in different reference frames.
Proc. Natl. Acad. Sci. USA, 108-28:11668-11673, 2011
Funded by: NWO Earth
and Life Sciences
Back to top, to Jeroen's Homepage, or to the Department’s homepage
10.
Representation of
Head-centric flow in the human motion complex
Research group: J. Goossens, S. Dukelow, R. Menon (Robarts Institute, London, ON), T. Vilis (UWO, London, ON) and A.V. van den Berg (Univ. Utrecht),
Summary
Recent neuro-imaging studies have identified putative homologues of macaque areas MT and MST in humans. Little is known about the integration of visual and non-visual signals in human motion areas compared to monkeys. Through extra-retinal signals the brain can factor out the components of visual flow on the retina that are induced by eye-in-head and head-in-space rotations, and achieve a representation of flow relative to the head (head-centric flow) or body (body-centric flow). Here, we used fMRI to test whether extraretinal eye-movement signals modulate responses to visual flow in the human MT+ complex. We distinguished between MT and MST, and tested whether subdivisions of these areas may transform the retinal flow into head-centric flow. We report that interactions between eye movement signals and visual flow are not evenly distributed across MT+. Pursuit hardly influenced the response of MT to flow while the responses in MST to the same retinal stimuli were stronger during pursuit than during fixation. We also identified two sub regions in which the flow-related responses were boosted significantly by pursuit, one overlapping part of MST. In addition, we find evidence of a metric relation between rotational flow relative to the head and fMRI signals in a sub region of MST. The latter findings provide an important advance over published single-cell recordings in monkey MST. A visual representation of the head’s rotation in the world derived from head-centric flow may supplement semicircular canals signals, and is appropriate for cross-calibrating vestibular and visual signals.
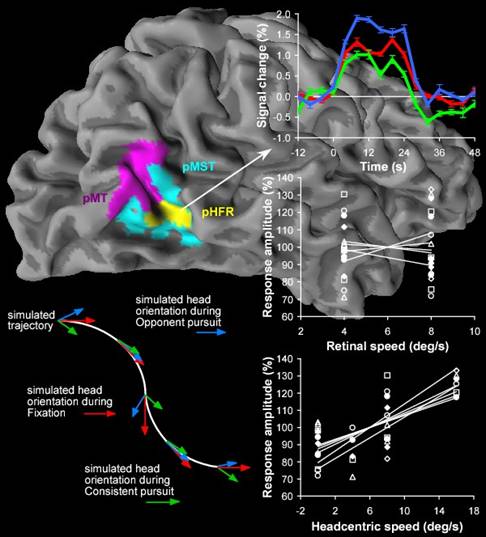
Publications
o
H.H.L.M. Goossens,
S.P. Dukelow, R.S. Menon, T. Vilis. A.V. van den Berg,
Representation
of head-centric flow in the human motion complex.
J. Neurosci.,
26:5616-5627, 2006.
o H.C. Golz, S.P. Dukelow, J.F.X. DeSouza, J.C. Culham, A.V. van den Berg, H.H.L.M. Goossens, R.S. Menon, T. Vilis.
A
putative homologue of monkey area VIP in humans.
Soc. Neurosci. Abstr, 27, 58.3, 2001